
 |
| https://vignette.wikia.nocookie.net/dinosaurrevolution/images/d/da/Troodon.jpg/revision/latest?cb=20130611200612 |
4) Troodon, Oviraptor, & Citipati in particular & non-bird maniraptorans in general had paternal care, "possibly within a polygamous mating system" ( http://www.esf.edu/EFB/faculty/documents/varricchio2008paternalcaredinosours.pdf ). I have 2 major problems w/this hypothesis: 1) Varricchio et al. used maximum rather than average values for clutch size (which didn't make sense to me given that "Troodon clutches vary from 12 to 24 eggs": http://studentresearch.wcp.muohio.edu/vertebrateevolution/dinoasurreprodparenting00.pdf ); 2) Varricchio et al. sampled only 2 oviraptorids (which didn't make sense to me given that "multiple brooding oviraptorids are known": http://dml.cmnh.org/2011Jan/msg00260.html ).

{kind=link}
{kind=link}
3) Dracorex & Stygimoloch were juvenile & sub-adult stages, respectively, of Pachycephalosaurus ( http://www.plosone.org/article/info:doi%2F10.1371%2Fjournal.pone.0007626 ). I have 2 major problems w/this hypothesis: 1) It's based on a CAST of the Dracorex skull, & thus it ignores the adult features of the skull (See "DISCUSSION": http://www.childrensmuseum.org/themuseum/dinosphere/draco_rex/dracorex_hogwartsia.pdf ); 2) It ignores the non-ontogenetic differences btwn Dracorex & Stygimoloch (Again, see "DISCUSSION": http://www.childrensmuseum.org/themuseum/dinosphere/draco_rex/dracorex_hogwartsia.pdf ).
 |
| https://luisvrey.files.wordpress.com/2012/06/deinob22.jpg?w=640 |
{kind=link}
 |
| https://i.infopls.com/images/EEX_DIN054FEASAU_001.jpg |
{kind=link}
1) Sauropods lacked an avian-style gastric mill & compensated for this "by greatly increasing food retention time in the digestive system" ( http://rspb.royalsocietypublishing.org/content/274/1610/635.full.pdf ). I have 2 major problems w/this hypothesis: 1) It's based on the comparison of sauropod gastroliths to ostrich gastroliths while ignoring moa gastroliths; Like sauropods, moas were herbivorous browsers (See the Shugart quote) while ostriches are omnivorous grazers ( http://wildwelfare.org/wp-content/uploads/2017/09/Common-Ostrich.pdf ); It's probably no coincidence, then, that like sauropod gastroliths, moa gastroliths are polished (Again, see the Shugart quote) while ostrich gastroliths are pitted; 2) It fails to explain sauropod digestion for the same reason that gigantothermy fails to explain non-bird dino physiology; To quote Holtz, it "might apply to large dinosaurs, but would not apply to small species or to babies" (See "Gigantothermy" under "Complications": http://www.geol.umd.edu/~tholtz/G104/lectures/104endo.html ).
 |
| https://i.ytimg.com/vi/midGaNNHhbM/hqdefault.jpg |
*E.g. It's implied that lone adult Komodo dragons can kill prey 10x their size w/"only ser- rated teeth", the logic being that lone adult Deinonychus would've done the same. However, it's been known since 2005 that the former are venomous ( http://www.academia.edu/462746/Early_evolution_of_the_venom_system_in_lizards_and_snakes ), hence why they can kill prey 10x their size. It's also implied, based on Horner & Dobb 1997, that the multiple Deinonychus individuals represented at YPM 64-75 were immature, the logic being that "larger (older) animals are more voracious cannibals than smaller (younger) animals, and smaller conspecifics are more often eaten than larger". However, Horner & Dobb 1997 is neither a peer-reviewed source nor points to a peer-reviewed source, & thus "the information is not likely to be useful" ( http://anthropology.ua.edu/bindon/ant570/pap_rule.htm ). AFAIK, the only relevant peer-reviewed source is Ostrom 1969 (according to which there is no "evidence of immature individuals at this site": http://peabody.yale.edu/sites/default/files/documents/scientific-publications/ypmB30_1969.pdf ).
Quoting Maxwell ( http://www.naturalhistorymag.com/htmlsite/master.html?http://www.naturalhistorymag.com/htmlsite/1299/1299_feature.html ): "Laid out in its death pose at this new site was a beautifully preserved, near-complete specimen of a young Tenontosaurus. Four Deinonychus teeth were found alongside the bones; later, in the laboratory, seven more teeth were uncovered. It's possible that a few more teeth were missed in the field or unwittingly discarded during preparation because they were concealed within small lumps of rock. So we have a subadult Tenontosaurus no more than fourteen feet long (compared with a length of about twenty feet for the adult at the Shrine site), preserved with at least eleven Deinonychus teeth.
But how can we distinguish between the remains of a victim hunted down and devoured by a pack and an animal that simply died and was scavenged by a few passing Deinonychus? As is the case at the Shrine site, this Tenontosaurus was preserved where it died. After death, the desiccation of the abundant supporting tendons that line the vertebrae of the neck and tail cause these parts to coil. The tail of Tenontosaurus, which accounts for about one-third of the animal's total length, is particularly heavy with supporting tendons. In this specimen, the pronounced curvature of the tail and the neck toward each other effectively counters any claim that the bones were carried to the site by water currents. The Deinonychus teeth were found in the region of the abdomen and pelvis, suggesting that the predators lost their teeth while feeding on the viscera. Most modern carnivores begin with the areas around the anus and abdomen when they feast on freshly killed prey, and it's likely that carnivorous dinosaurs did the same.
The number of teeth indicate that more than one Deinonychus was involved with the carcass. Like all other theropod dinosaurs, Deinonychus shed and replaced teeth throughout its life. The teeth would fall out upon the animal's reaching maturity but also could be wrenched out earlier by the stress associated with the biting and tearing of flesh. Because of this, theropod teeth are very common in sediments containing dinosaur fossils. The teeth from this site vary from recently erupted to fully mature ones. Given that Deinonychus had only sixty teeth in its jaws at any one time, it's unlikely that all eleven were wrenched from the mouth of just one feeding animal. This would leave the Deinonychus toothless after five similar meals. The possibility that Deinonychus was replacing shed teeth in a few weeks or months, and therefore had the ability to sustain such dramatic tooth loss, was quashed by Greg Erickson, who, as a master's degree student at the Museum of the Rockies, worked on replacement rates of teeth in various dinosaurs and living reptiles. After CT-scanning portions of the lower jaw of Deinonychus and studying individual teeth, he came up with an estimate of 300 days for the time it took Deinonychus to replace a shed tooth with a mature one.
We know that this Tenontosaurus was not yet an adult, so it didn't die of old age. Of course, this doesn't rule out death from disease or injury and doesn't confirm that it was cut down by a pack, but it's a start. Next, we have a concentration of teeth around the abdomen and pelvis. This may indicate that the pack fed on the abdominal contents while they were still warm and moist. If, after the viscera had been consumed, the remainder of the carcass was scavenged over time by many individuals, we would expect a much more disturbed carcass and a wider scattering of teeth. Similarly, if the Tenontosaurus had been killed by a larger predator-such as the unknown owner of the three-inch-long serrated teeth that occasionally crop up in the Cloverly formation—then whatever remained of the carcass would have been strewn around the area."
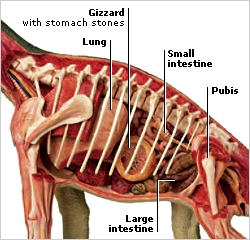
Quoting Shugart ( http://www.amazon.com/Earthquake-Other-Tales-Unbalanced-Nature/dp/0300122705 ): "When the Polynesians arrived in New Zealand, they encountered birds that had been evolving for 80 million years without the presence of mammalian predators. Among the most striking of these animals must have been the moas (Figure 29). These were gigantic wingless birds standing as much as 10 feet (3 m) tall and weighing as much as 550 pounds (250 kg).1 They are known from a diverse array of remains including eggshells, eggs, a few mummified carcasses, vast numbers of bones, and some older fossilized bone. The eleven moa species that are currently recognized occupied ecological niches customarily filled elsewhere by large mammalian browsing herbivores. They may have had relatively low reproductive rates; apparently, they usually laid only one egg at a time.2
Moas ranked in height from the tallest at about 10 feet to smaller species the size of a large domestic turkey (about 3 feet, or 1 m, tall and weighing 45 pounds, or 20 kg). They were unique in having neither wings nor even residual wing bones. As one expects of large birds that feed on vegetable matter, moas had muscular gizzards. They swallowed small stones up to 2 inches (50 mm) in diameter into their gizzards for grinding food before digestion. These polished stones, called gastroliths, often occur in groups along with moa bones.3
Many gastroliths have been found in what are now human-modified grassy habitats, giving the initial impression that moas were grass-eating animals. But the present vegetation at a site may not be its previous vegetation.4 Based on preserved crop contents from mummified specimens, moas fed on leaves, seeds, and green twigs of trees and parts of shrubs.5 Thus, it appears that they were creatures of the forest and shrub- land—more like browsing deer than grazing cows."